
In molecular biology, mutagenesis is an important laboratory technique whereby DNA mutations are deliberately engineered to produce libraries of mutant genes, proteins, strains of bacteria, or other genetically modified organisms. The various constituents of a gene, as well as its regulatory elements and its gene products, may be mutated so that the functioning of a genetic locus, process, or product can be examined in detail. The mutation may produce mutant proteins with interesting properties or enhanced or novel functions that may be of commercial use. Mutant strains may also be produced that have practical application or allow the molecular basis of a particular cell function to be investigated.
Many methods of mutagenesis exist today. Initially, the kind of mutations artificially induced in the laboratory were entirely random using mechanisms such as UV irradiation. Random mutagenesis cannot target specific regions or sequences of the genome; however, with the development of site-directed mutagenesis, more specific changes can be made. Since 2013, development of the CRISPR/Cas9 technology, based on a prokaryotic viral defense system, has allowed for the editing or mutagenesis of a genome in vivo.[1] Site-directed mutagenesis has proved useful in situations that random mutagenesis is not. Other techniques of mutagenesis include combinatorial and insertional mutagenesis. Mutagenesis that is not random can be used to clone DNA,[2] investigate the effects of mutagens,[3] and engineer proteins.[4] It also has medical applications such as helping immunocompromised patients, research and treatment of diseases including HIV and cancers, and curing of diseases such as beta thalassemia.[5]
Random mutagenesis
Early approaches to mutagenesis relied on methods which produced entirely random mutations. In such methods, cells or organisms are exposed to mutagens such as UV radiation or mutagenic chemicals, and mutants with desired characteristics are then selected. Hermann Muller discovered in 1927 that X-rays can cause genetic mutations in fruit flies,[6] and went on to use the mutants he created for his studies in genetics.[7] For Escherichia coli, mutants may be selected first by exposure to UV radiation, then plated onto an agar medium. The colonies formed are then replica-plated, one in a rich medium, another in a minimal medium, and mutants that have specific nutritional requirements can then be identified by their inability to grow in the minimal medium. Similar procedures may be repeated with other types of cells and with different media for selection.
A number of methods for generating random mutations in specific proteins were later developed to screen for mutants with interesting or improved properties. These methods may involve the use of doped nucleotides in oligonucleotide synthesis, or conducting a PCR reaction in conditions that enhance misincorporation of nucleotides (error-prone PCR), for example by reducing the fidelity of replication or using nucleotide analogues.[8] A variation of this method for integrating non-biased mutations in a gene is sequence saturation mutagenesis.[9] PCR products which contain mutation(s) are then cloned into an expression vector and the mutant proteins produced can then be characterised.
In animal studies, alkylating agents such as N-ethyl-N-nitrosourea (ENU) have been used to generate mutant mice.[10][11] Ethyl methanesulfonate (EMS) is also often used to generate animal, plant, and virus mutants.[12][13][14]
In a European Union law (as 2001/18 directive), this kind of mutagenesis may be used to produce GMOs but the products are exempted from regulation: no labeling, no evaluation.[15]
Site-directed mutagenesis
Prior to the development site-directed mutagenesis techniques, all mutations made were random, and scientists had to use selection for the desired phenotype to find the desired mutation. Random mutagenesis techniques has an advantage in terms of how many mutations can be produced; however, while random mutagenesis can produce a change in single nucleotides, it does not offer much control as to which nucleotide is being changed.[5] Many researchers therefore seek to introduce selected changes to DNA in a precise, site-specific manner. Early attempts uses analogs of nucleotides and other chemicals were first used to generate localized point mutations.[16] Such chemicals include aminopurine, which induces an AT to GC transition,[17] while nitrosoguanidine,[18] bisulfite,[19] and N4-hydroxycytidine may induce a GC to AT transition.[20][21] These techniques allow specific mutations to be engineered into a protein; however, they are not flexible with respect to the kinds of mutants generated, nor are they as specific as later methods of site-directed mutagenesis and therefore have some degree of randomness. Other technologies such as cleavage of DNA at specific sites on the chromosome, addition of new nucleotides, and exchanging of base pairs it is now possible to decide where mutations can go.[11][8]
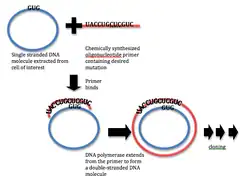
Current techniques for site-specific mutation originates from the primer extension technique developed in 1978. Such techniques commonly involve using pre-fabricated mutagenic oligonucleotides in a primer extension reaction with DNA polymerase. This methods allows for point mutation or deletion or insertion of small stretches of DNA at specific sites. Advances in methodology have made such mutagenesis now a relatively simple and efficient process.[3]
Newer and more efficient methods of site directed mutagenesis are being constantly developed. For example, a technique called "Seamless ligation cloning extract" (or SLiCE for short) allows for the cloning of certain sequences of DNA within the genome, and more than one DNA fragment can be inserted into the genome at once.[2]
Site directed mutagenesis allows the effect of specific mutation to be investigated. There are numerous uses; for example, it has been used to determine how susceptible certain species were to chemicals that are often used In labs. The experiment used site directed mutagenesis to mimic the expected mutations of the specific chemical. The mutation resulted in a change in specific amino acids and the effects of this mutation were analyzed.[3]

The site-directed approach may be done systematically in such techniques as alanine scanning mutagenesis, whereby residues are systematically mutated to alanine in order to identify residues important to the structure or function of a protein.[22] Another comprehensive approach is site saturation mutagenesis where one codon or a set of codons may be substituted with all possible amino acids at the specific positions.[23][24]
Combinatorial mutagenesis
Combinatorial mutagenesis is a site-directed protein engineering technique whereby multiple mutants of a protein can be simultaneously engineered based on analysis of the effects of additive individual mutations.[25] It provides a useful method to assess the combinatorial effect of a large number of mutations on protein function.[26] Large numbers of mutants may be screened for a particular characteristic by combinatorial analysis.[25] In this technique, multiple positions or short sequences along a DNA strand may be exhaustively modified to obtain a comprehensive library of mutant proteins.[25] The rate of incidence of beneficial variants can be improved by different methods for constructing mutagenesis libraries. One approach to this technique is to extract and replace a portion of the DNA sequence with a library of sequences containing all possible combinations at the desired mutation site. The content of the inserted segment can include sequences of structural significance, immunogenic property, or enzymatic function. A segment may also be inserted randomly into the gene in order to assess structural or functional significance of a particular part of a protein.[25]
Insertional mutagenesis
The insertion of one or more base pairs, resulting in DNA mutations, is also known as insertional mutagenesis.[27] Engineered mutations such as these can provide important information in cancer research, such as mechanistic insights into the development of the disease. Retroviruses and transposons are the chief instrumental tools in insertional mutagenesis. Retroviruses, such as the mouse mammory tumor virus and murine leukemia virus, can be used to identify genes involved in carcinogenesis and understand the biological pathways of specific cancers.[28] Transposons, chromosomal segments that can undergo transposition, can be designed and applied to insertional mutagenesis as an instrument for cancer gene discovery.[28] These chromosomal segments allow insertional mutagenesis to be applied to virtually any tissue of choice while also allowing for more comprehensive, unbiased depth in DNA sequencing.[28]
Researchers have found four mechanisms of insertional mutagenesis that can be used on humans. the first mechanism is called enhancer insertion. Enhancers boost transcription of a particular gene by interacting with a promoter of that gene. This particular mechanism was first used to help severely immunocompromised patients I need of bone marrow. Gammaretroviruses carrying enhancers were then inserted into patients. The second mechanism is referred to as promoter insertion. Promoters provide our cells with the specific sequences needed to begin translation. Promoter insertion has helped researchers learn more about the HIV virus. The third mechanism is gene inactivation. An example of gene inactivation is using insertional mutagenesis to insert a retrovirus that disrupts the genome of the T cell in leukemia patients and giving them a specific antigen called CAR allowing the T cells to target cancer cells. The final mechanisms is referred to as mRNA 3' end substitution. Our genes occasionally undergo point mutations causing beta-thalassemia that interrupts red blood cell function. To fix this problem the correct gene sequence for the red blood cells are introduced and a substitution is made.[5]
Homologous recombination
Homologous recombination can be used to produce specific mutation in an organism. Vector containing DNA sequence similar to the gene to be modified is introduced to the cell, and by a process of recombination replaces the target gene in the chromosome. This method can be used to introduce a mutation or knock out a gene, for example as used in the production of knockout mice.[29]
CRISPR
Since 2013, the development of CRISPR-Cas9 technology has allowed for the efficient introduction of different types of mutations into the genome of a wide variety of organisms. The method does not require a transposon insertion site, leaves no marker, and its efficiency and simplicity has made it the preferred method for genome editing.[30][31]
Gene synthesis
As the cost of DNA oligonucleotide synthesis falls, artificial synthesis of a complete gene is now a viable method for introducing mutations into a gene. This method allows for extensive mutation at multiple sites, including the complete redesign of the codon usage of a gene to optimise it for a particular organism.[32]
See also
References
- ↑ Hsu PD, Lander ES, Zhang F (June 2014). "Development and applications of CRISPR-Cas9 for genome engineering". Cell. 157 (6): 1262–78. doi:10.1016/j.cell.2014.05.010. PMC 4343198. PMID 24906146.
- 1 2 Motohashi K (June 2015). "A simple and efficient seamless DNA cloning method using SLiCE from Escherichia coli laboratory strains and its application to SLiP site-directed mutagenesis". BMC Biotechnology. 15: 47. doi:10.1186/s12896-015-0162-8. PMC 4453199. PMID 26037246.
- 1 2 3 Doering JA, Lee S, Kristiansen K, Evenseth L, Barron MG, Sylte I, LaLone CA (November 2018). "In Silico Site-Directed Mutagenesis Informs Species-Specific Predictions of Chemical Susceptibility Derived From the Sequence Alignment to Predict Across Species Susceptibility (SeqAPASS) Tool". Toxicological Sciences. 166 (1): 131–145. doi:10.1093/toxsci/kfy186. PMC 6390969. PMID 30060110.
- ↑ Choi GC, Zhou P, Yuen CT, Chan BK, Xu F, Bao S, et al. (August 2019). "Combinatorial mutagenesis en masse optimizes the genome editing activities of SpCas9". Nature Methods. 16 (8): 722–730. doi:10.1038/s41592-019-0473-0. PMID 31308554. S2CID 196811756.
- 1 2 3 Bushman FD (February 2020). "Retroviral Insertional Mutagenesis in Humans: Evidence for Four Genetic Mechanisms Promoting Expansion of Cell Clones". Molecular Therapy. 28 (2): 352–356. doi:10.1016/j.ymthe.2019.12.009. PMC 7001082. PMID 31951833.
- ↑ Muller HJ (July 1927). "Artificial Transmutation of the Gene" (PDF). Science. 66 (1699): 84–7. Bibcode:1927Sci....66...84M. doi:10.1126/science.66.1699.84. PMID 17802387.
- ↑ Crow JF, Abrahamson S (December 1997). "Seventy years ago: mutation becomes experimental". Genetics. 147 (4): 1491–6. doi:10.1093/genetics/147.4.1491. PMC 1208325. PMID 9409815.
- 1 2 Blackburn GM, ed. (2006). Nucleic Acids in Chemistry and Biology (3rd ed.). Royal Society of Chemistry. pp. 191–192. ISBN 978-0854046546.
- ↑ Wong TS, Tee KL, Hauer B, Schwaneberg U (February 2004). "Sequence saturation mutagenesis (SeSaM): a novel method for directed evolution". Nucleic Acids Research. 32 (3): 26e–26. doi:10.1093/nar/gnh028. PMC 373423. PMID 14872057.
- ↑ Justice MJ, Noveroske JK, Weber JS, Zheng B, Bradley A (1999). "Mouse ENU mutagenesis". Human Molecular Genetics. 8 (10): 1955–63. doi:10.1093/hmg/8.10.1955. PMID 10469849.
- 1 2 Hrabé de Angelis M, Balling R (May 1998). "Large scale ENU screens in the mouse: genetics meets genomics". Mutation Research. 400 (1–2): 25–32. doi:10.1016/s0027-5107(98)00061-x. PMID 9685575.
- ↑ Flibotte S, Edgley ML, Chaudhry I, Taylor J, Neil SE, Rogula A, et al. (June 2010). "Whole-genome profiling of mutagenesis in Caenorhabditis elegans". Genetics. 185 (2): 431–41. doi:10.1534/genetics.110.116616. PMC 2881127. PMID 20439774.
- ↑ Bökel C (2008). EMS screens : from mutagenesis to screening and mapping. Methods in Molecular Biology. Vol. 420. pp. 119–38. doi:10.1007/978-1-59745-583-1_7. PMID 18641944.
- ↑ Favor AH, Llanos CD, Youngblut MD, Bardales JA (2020). "Optimizing bacteriophage engineering through an accelerated evolution platform". Scientific Reports. 10 (1): 13981. doi:10.1038/s41598-020-70841-1. PMC 7438504. PMID 32814789.
- ↑ Krinke C (March 2018). "GMO directive : the origins of the mutagenesis exemption". Inf'OGM.
- ↑ Shortle D, DiMaio D, Nathans D (1981). "Directed mutagenesis". Annual Review of Genetics. 15: 265–94. doi:10.1146/annurev.ge.15.120181.001405. PMID 6279018.
- ↑ Caras IW, MacInnes MA, Persing DH, Coffino P, Martin DW (September 1982). "Mechanism of 2-aminopurine mutagenesis in mouse T-lymphosarcoma cells". Molecular and Cellular Biology. 2 (9): 1096–103. doi:10.1128/mcb.2.9.1096. PMC 369902. PMID 6983647.
- ↑ McHugh GL, Miller CG (October 1974). "Isolation and characterization of proline peptidase mutants of Salmonella typhimurium". Journal of Bacteriology. 120 (1): 364–71. doi:10.1128/JB.120.1.364-371.1974. PMC 245771. PMID 4607625.
- ↑ Shortle D, Nathans D (May 1978). "Local mutagenesis: a method for generating viral mutants with base substitutions in preselected regions of the viral genome". Proceedings of the National Academy of Sciences of the United States of America. 75 (5): 2170–4. Bibcode:1978PNAS...75.2170S. doi:10.1073/pnas.75.5.2170. PMC 392513. PMID 209457.
- ↑ Flavell RA, Sabo DL, Bandle EF, Weissmann C (January 1975). "Site-directed mutagenesis: effect of an extracistronic mutation on the in vitro propagation of bacteriophage Qbeta RNA". Proceedings of the National Academy of Sciences of the United States of America. 72 (1): 367–71. Bibcode:1975PNAS...72..367F. doi:10.1073/pnas.72.1.367. PMC 432306. PMID 47176.
- ↑ Müller W, Weber H, Meyer F, Weissmann C (September 1978). "Site-directed mutagenesis in DNA: generation of point mutations in cloned beta globin complementary dna at the positions corresponding to amino acids 121 to 123". Journal of Molecular Biology. 124 (2): 343–58. doi:10.1016/0022-2836(78)90303-0. PMID 712841.
- ↑ Vanessa E. Gray; Ronald J. Hause; Douglas M. Fowler (September 1, 2017). "Analysis of Large-Scale Mutagenesis Data To Assess the Impact of Single Amino Acid Substitutions". Genetics. 207 (1): 53–61. doi:10.1534/genetics.117.300064. PMC 5586385. PMID 28751422.
- ↑ Reetz, M. T.; Carballeira J. D. (2007). "Iterative saturation mutagenesis (ISM) for rapid directed evolution of functional enzymes". Nature Protocols. 2 (4): 891–903. doi:10.1038/nprot.2007.72. PMID 17446890. S2CID 37361631.
- ↑ Cerchione, Derek; Loveluck, Katherine; Tillotson, Eric L.; Harbinski, Fred; DaSilva, Jen; Kelley, Chase P.; Keston-Smith, Elise; Fernandez, Cecilia A.; Myer, Vic E.; Jayaram, Hariharan; Steinberg, Barrett E.; Xu, Shuang-yong (16 April 2020). "SMOOT libraries and phage-induced directed evolution of Cas9 to engineer reduced off-target activity". PLOS ONE. 15 (4): e0231716. Bibcode:2020PLoSO..1531716C. doi:10.1371/journal.pone.0231716. PMC 7161989. PMID 32298334.
- 1 2 3 4 Parker AS, Griswold KE, Bailey-Kellogg C (November 2011). "Optimization of combinatorial mutagenesis". Journal of Computational Biology. 18 (11): 1743–56. Bibcode:2011LNCS.6577..321P. doi:10.1089/cmb.2011.0152. PMC 5220575. PMID 21923411.
- ↑ Choi GC, Zhou P, Yuen CT, Chan BK, Xu F, Bao S, Chu HY, Thean D, Tan K, Wong KH, Zheng Z, Wong AS (August 2019). "Combinatorial mutagenesis en masse optimizes the genome editing activities of SpCas9". Nature Methods. 16 (8): 722–730. doi:10.1038/s41592-019-0473-0. PMID 31308554. S2CID 196811756.
- ↑ Uren AG, Kool J, Berns A, van Lohuizen M (November 2005). "Retroviral insertional mutagenesis: past, present and future". Oncogene. 24 (52): 7656–72. doi:10.1038/sj.onc.1209043. PMID 16299527. S2CID 14441244.
- 1 2 3 Vassiliou G, Rad R, Bradley A (2010-01-01). "The use of DNA transposons for cancer gene discovery in mice". In Wassarman PM, Soriano PM (eds.). Guide to Techniques in Mouse Development, Part B: Mouse Molecular Genetics, 2nd Edition. Methods in Enzymology. Vol. 477 (2nd ed.). Academic Press. pp. 91–106. doi:10.1016/s0076-6879(10)77006-3. ISBN 9780123848802. PMID 20699138.
- ↑ "Homologous Recombination Method (and Knockout Mouse)". Davidson College.
- ↑ Damien Biot-Pelletier; Vincent J. J. Martin (2016). "Seamless site-directed mutagenesis of the Saccharomyces cerevisiae genome using CRISPR-Cas9". Journal of Biological Engineering. 10: 6. doi:10.1186/s13036-016-0028-1. PMC 4850645. PMID 27134651.
- ↑ Xu S (20 August 2015). "The application of CRISPR-Cas9 genome editing in Caenorhabditis elegans". J Genet Genomics. 42 (8): 413–21. doi:10.1016/j.jgg.2015.06.005. PMC 4560834. PMID 26336798.
- ↑ Khudyakov YE, Fields HA, eds. (25 September 2002). Artificial DNA: Methods and Applications. CRC Press. p. 13. ISBN 9781420040166.
External links
| Library resources about Mutagenesis |