| Argentinosaurus | |
|---|---|
 | |
| Reconstructed skeleton, Museo Municipal Carmen Funes, Plaza Huincul, Argentina. The original vertebrae are seen on the lower left | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | †Sauropodomorpha |
| Clade: | †Sauropoda |
| Clade: | †Macronaria |
| Clade: | †Titanosauria |
| Clade: | †Lognkosauria |
| Genus: | †Argentinosaurus Bonaparte & Coria, 1993 |
| Type species | |
| †Argentinosaurus huinculensis Bonaparte & Coria, 1993 | |
Argentinosaurus is a genus of giant sauropod dinosaur that lived during the Late Cretaceous period in what is now Argentina. Although it is only known from fragmentary remains, Argentinosaurus is one of the largest known land animals of all time, perhaps the largest, measuring 30–35 metres (98–115 ft) long and weighing 65–80 tonnes (72–88 short tons). It was a member of Titanosauria, the dominant group of sauropods during the Cretaceous. It is widely regarded by many paleontologists as the biggest dinosaur ever, and perhaps lengthwise the longest animal ever, though both claims have no concrete evidence yet.
The first Argentinosaurus bone was discovered in 1987 by a farmer on his farm near the city of Plaza Huincul. A scientific excavation of the site led by the Argentine palaeontologist José Bonaparte was conducted in 1989, yielding several back vertebrae and parts of a sacrum—fused vertebrae between the back and tail vertebrae. Additional specimens include a complete femur (thigh bone) and the shaft of another. Argentinosaurus was named by Bonaparte and the Argentine palaeontologist Rodolfo Coria in 1993; the genus contains a single species, A. huinculensis. The generic name Argentinosaurus means "Argentine lizard", and the specific name huinculensis refers to its place of discovery, Plaza Huincul.
The fragmentary nature of Argentinosaurus remains makes their interpretation difficult. Arguments revolve around the position of the recovered vertebrae within the vertebral column and the presence of accessory articulations between the vertebrae that would have strengthened the spine. A computer model of the skeleton and muscles estimated this dinosaur had a maximum speed of 7 km/h (5 mph) with a pace, a gait where the fore and hind limb of the same side of the body move simultaneously. The fossils of Argentinosaurus were recovered from the Huincul Formation, which was deposited in the middle Cenomanian to early Turonian ages (about 96 to 92 million years ago) and contains a diverse dinosaur fauna including the giant theropod Mapusaurus.
Discovery

The first Argentinosaurus bone, which is now thought to be a fibula (calf bone), was discovered in 1987 by Guillermo Heredia on his farm "Las Overas" about 8 km (5 mi) east of Plaza Huincul, in Neuquén Province, Argentina. Heredia, initially believing he had discovered petrified logs, informed the local museum, the Museo Carmen Funes, whose staff members excavated the bone and stored it in the museum's exhibition room. In early 1989, the Argentine palaeontologist José F. Bonaparte initiated a larger excavation of the site involving palaeontologists of the Museo Argentino de Ciencias Naturales, yielding a number of additional elements from the same individual. The individual, which later became the holotype of Argentinosaurus huinculensis, is catalogued under the specimen number MCF-PVPH 1.[1]
Separating fossils from the very hard rock in which the bones were encased required the use of pneumatic hammers.[2][3][4]: 35 The additional material recovered included seven dorsal vertebrae (vertebrae of the back),[1] the underside of the sacrum (fused vertebrae between the dorsal and tail vertebrae) including the first to fifth sacral vertebrae and some sacral ribs, and a part of a dorsal rib (rib from the flank).[2] These finds were also incorporated into the collection of the Museo Carmen Funes.[2]
Bonaparte presented the new find in 1989 at a scientific conference in San Juan. The formal description was published in 1993 by Bonaparte and the Argentine palaeontologist Rodolfo Coria, with the naming of a new genus and species, Argentinosaurus huinculensis. The generic name means "Argentine lizard", while the specific name refers to the town Plaza Huincul.[2] Bonaparte and Coria described the limb bone discovered in 1987 as an eroded tibia (shin bone), although the Uruguayan palaeontologist Gerardo Mazzetta and colleagues reidentified this bone as a left fibula in 2004.[5][6] In 1996, Bonaparte referred (assigned) a complete femur (thigh bone) from the same locality to the genus, which was put on exhibit at the Museo Carmen Funes. This bone was deformed by front-to-back crushing during fossilization. In their 2004 study, Mazzetta and colleagues mentioned an additional femur that is housed in the La Plata Museum under the specimen number MLP-DP 46-VIII-21-3. Though not as strongly deformed as the complete femur, it preserves only the shaft and lacks its upper and lower ends. Both specimens belonged to individuals equivalent in size to the holotype individual.[5] As of 2019, however, it was still uncertain whether any of these femora belonged to Argentinosaurus.[7]
Description
Size
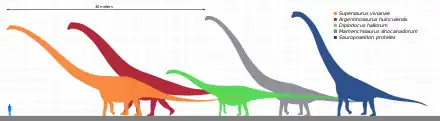
Argentinosaurus is among the largest known land animals, although its exact size is difficult to estimate because of the incompleteness of its remains.[8] To counter this problem, palaeontologists can compare the known material with that of smaller related sauropods known from more complete remains. The more complete taxon can then be scaled up to match the dimensions of Argentinosaurus. Mass can be estimated from known relationships between certain bone measurements and body mass, or through determining the volume of models.[9]
A reconstruction of Argentinosaurus created by Gregory Paul in 1994 yielded a length estimate of 30–35 metres (98–115 ft).[10] Later that year, estimates by Bonaparte and Coria suggesting a hind limb length of 4.5 metres (15 ft), a trunk length (hip to shoulder) of 7 metres (23 ft), and an overall body length of 30 metres (98 ft) were published.[11] In 2006, Kenneth Carpenter reconstructed Argentinosaurus using the more complete Saltasaurus as a guide and estimated a length of 30 metres (98 ft).[12] In 2008, Jorge Calvo and colleagues used the proportions of Futalognkosaurus to estimate the length of Argentinosaurus at less than 33 metres (108 ft).[13] In 2013, William Sellers and colleagues arrived at a length estimate of 39.7 metres (130 ft) and a shoulder height of 7.3 metres (24 ft) by measuring the skeletal mount in Museo Carmen Funes.[14] During the same year, Scott Hartman suggested that because Argentinosaurus was then thought to be a basal titanosaur, it would have a shorter tail and narrower chest than Puertasaurus, which he estimated to be about 27 metres (89 ft) long, indicating Argentinosaurus was slightly smaller.[15] In 2016, Paul estimated the length of Argentinosaurus at 30 m (98 ft),[16] but later estimated a greater length of 35 metres (115 ft) or longer in 2019, restoring the unknown neck and tail of Argentinosaurus after those of other large South American titanosaurs.[7]

Paul estimated a body mass of 80–100 tonnes (88–110 short tons) for Argentinosaurus in 1994.[10] In 2004, Mazzetta and colleagues provided a range of 60–88 tonnes (66–97 short tons) and considered 73 tonnes (80 short tons) to be the most likely mass, making it the heaviest sauropod known from good material.[5] In 2013, Sellers and colleagues estimated a mass of 83.2 tonnes (91.7 short tons) by calculating the volume of the aforementioned Museo Carmen Funes skeleton.[14] In 2014 and 2018, Roger Benson and colleagues estimated the mass of Argentinosaurus at 90 and 95 tonnes (99 and 105 short tons),[17][18] but these estimates were questioned due to a very large error range and lack of precision.[19] In 2016, using equations that estimate body mass based on the circumference of the humerus and femur of quadrupedal animals, Bernardo Gonzáles Riga and colleagues estimated a mass of 96.4 tonnes (106.3 short tons) based on an isolated femur; it is uncertain whether this femur actually belongs to Argentinosaurus.[20] In the same year, Paul moderated his earlier estimate from 1994 and listed the body mass of Argentinosaurus at more than 50 tonnes (55 short tons).[16] In 2019, Paul moderated his 2016 estimate and gave a mass estimate of 65–75 tonnes (72–83 short tons) based on his skeletal reconstructions (diagrams illustrating the bones and shape of an animal) of Argentinosaurus in dorsal and lateral view.[7] In 2020, Campione and Evans also yielded a body mass estimate of approximately 75 tonnes (83 short tons).[19] In 2023, Paul and Larramendi proposed that the holotype would have weighed between 75–80 metric tons (83–88 short tons) at maximum. They further suggested that the enigmatic, fragmentary Bruhathkayosaurus possibly weighed more, between 110 and 130 tonnes (120 and 140 short tons).[21]
While Argentinosaurus was definitely a massive animal, there is disagreement over whether it was the largest known titanosaur. Puertasaurus, Futalognkosaurus, Dreadnoughtus, Paralititan, "Antarctosaurus" giganteus, and Alamosaurus have all been considered to be comparable in size with Argentinosaurus by some studies,[22][23] although others have found them to be notably smaller.[13][24][7] In 2017, Carballido and colleagues considered Argentinosaurus to be smaller than Patagotitan, since the latter had a greater area enclosed by the neural spine, diapophyses, and parapophyses of its anterior dorsal vertebrae.[8] However, Paul found Patagotitan to be smaller than Argentinosaurus in 2019, due to the latter's dorsal column being considerably longer. Even if Argentinosaurus was the largest-known titanosaur, other sauropods including Maraapunisaurus and a giant mamenchisaurid, may have been larger, although these are only known from very scant remains. Some diplodocids, such as Supersaurus and Diplodocus[25][7] may have exceeded Argentinosaurus in length despite being considerably less massive.[12][26] The mass of the blue whale, however, which can be greater than 150 tonnes (170 short tons),[27][28] still exceeds that of all known sauropods.[7]
Vertebrae

Argentinosaurus likely possessed 10 dorsal vertebrae, like other titanosaurs.[7] The vertebrae were enormous even for sauropods; one dorsal vertebra has a reconstructed height of 159 centimetres (63 in) and a width of 129 centimetres (51 in), and the vertebral centra are up to 57 centimetres (22 in) in width.[2] In 2019, Paul estimated the total length of the dorsal vertebral column at 447 centimetres (176 in) and the width of the pelvis at 0.6 times the combined length of the dorsal and sacral vertebral column.[7] The dorsals were opisthocoelous (concave at the rear) as in other macronarian sauropods.[2][6]: 205 The pleurocoels (excavations on the sides of the centra) were proportionally small and positioned in the front half of the centrum.[29]: 102 The vertebrae were internally lightened by a complex pattern of numerous air-filled chambers. Such camellate bone is, among sauropods, especially pronounced in the largest and longest-necked species.[30][31] In both the dorsal and sacral vertebrae, very large cavities measuring 4 to 6 centimetres (1.6 to 2.4 in) were present.[30] The dorsal ribs were tubular and cylindrical in shape, in contrast with other titanosaurs.[2][32]: 309 Bonaparte and Coria, in their 1993 description, noted the ribs were hollow, unlike those of many other sauropods, but later authors argued this hollowing could also have been due to erosion after the death of the individual.[6] Argentinosaurus, like many titanosaurs,[33] probably had six sacral vertebrae (those in the hip region), although the last one is not preserved. The centra of the second to fifth sacral vertebrae were much reduced in size and considerably smaller than the centrum of the first sacral. The sacral ribs curved downwards. The second sacral rib was larger than the other preserved sacral ribs, though the size of the first is unknown due to its incompleteness.[2]

Because of their incomplete preservation, the original position of the known dorsal vertebrae within the vertebral column is disputed. Dissenting configurations were suggested by Bonaparte and Coria in 1993; Fernando Novas and Martín Ezcurra in 2006; and Leonardo Salgado and Jaime Powell in 2010. One vertebra was interpreted by these studies as the first, fifth or third; and another vertebra as the second, tenth or eleventh, or ninth, respectively. A reasonably complete vertebra was found to be the third by the 1993 and 2006 studies, but the fourth by the 2010 study. Another vertebra was interpreted by the three studies as being part of the rear section of the dorsal vertebral column, as the fourth, or as the fifth, respectively. In 1993, two articulated (still connected) vertebrae were thought to be of the rear part of the dorsal column but are interpreted as the sixth and seventh vertebrae in the two later studies. The 2010 study mentioned another vertebra that was not mentioned by the 1993 and 2006 studies; it was presumed to belong to the rear part of the dorsal column.[2][34][1]
Another contentious issue is the presence of hyposphene-hypantrum articulations, accessory joints between vertebrae that were located below the main articular processes. Difficulties in interpretation arise from the fragmentary preservation of the vertebral column; these joints are hidden from view in the two connected vertebrae.[30] In 1993, Bonaparte and Coria said the hyposphene-hypantrum articulations were enlarged, as in the related Epachthosaurus, and had additional articular surfaces that extended downwards.[2] This was confirmed by some later authors; Novas noted the hypantrum (a bony extension below the articular processes of the front face of a vertebra) extended sidewards and downwards, forming a much-broadened surface that connected with the equally enlarged hyposphene at the back face of the following vertebra.[30][32]: 309–310 In 1996, Bonaparte stated these features would have made the spine more rigid and were possibly an adaptation to the giant size of the animal.[29] Other authors argued most titanosaur genera lacked hyposphene-hypantrum articulations and that the articular structures seen in Epachthosaurus and Argentinosaurus are thickened vertebral laminae (ridges).[30][35][36]: 55 Sebastián Apesteguía, in 2005, argued the structures seen in Argentinosaurus, which he termed hyposphenal bars, are indeed thickened laminae that could have been derived from the original hyposphene and had the same function.[37]
Limbs
.jpg.webp)
The complete femur that was assigned to Argentinosaurus is 2.5 metres (8.2 ft) long. The femoral shaft has a circumference of about 1.18 metres (3.9 ft) at its narrowest part. Mazzetta and colleagues used regression equations to estimate its original length at 2.557 metres (8.39 ft), which is similar to the length of the other femur, and later in 2019 Paul gave a similar estimate of 2.575 metres (8.45 ft).[7] By comparison, the complete femora preserved in the other giant titanosaurs Antarctosaurus giganteus and Patagotitan mayorum measure 2.35 metres (7.7 ft) and 2.38 metres (7.8 ft), respectively.[5][8] While the holotype specimen does not preserve a femur, it preserves a slender fibula (originally interpreted as a tibia) that is 1.55 metres (5.1 ft) in length. When it was identified as a tibia, it was thought to have a comparatively short cnemial crest, a prominent extension at the upper front that anchored muscles for stretching the leg. However, as stated by Mazzetta and colleagues, this bone lacks both the proportions and anatomical details of a tibia, while being similar in shape to other sauropod fibulae.[2][5]
Classification
Relationships within Titanosauria are amongst the least understood of all groups of dinosaurs.[38] Traditionally, the majority of sauropod fossils from the Cretaceous had been referred to a single family, the Titanosauridae, which has been in use since 1893.[39] In their 1993 first description of Argentinosaurus, Bonaparte and Coria noted it differed from typical titanosaurids in having hyposphene-hypantrum articulations. As these articulations were also present in the titanosaurids Andesaurus and Epachthosaurus, Bonaparte and Coria proposed a separate family for the three genera, the Andesauridae. Both families were united into a new, higher group called Titanosauria.[2]
In 1997, Salgado and colleagues found Argentinosaurus to belong to Titanosauridae in an unnamed clade with Opisthocoelicaudia and an indeterminate titanosaur.[40] In 2002, Davide Pisani and colleagues recovered Argentinosaurus as a member of Titanosauria, and again found it to be in a clade with Opisthocoelicaudia and an unnamed taxon, in addition to Lirainosaurus.[41] A 2003 study by Jeffrey Wilson and Paul Upchurch found both Titanosauridae and Andesauridae to be invalid; the Titanosauridae because it was based on the dubious genus Titanosaurus and the Andesauridae because it was defined on plesiomorphies (primitive features) rather than on synapomorphies (newly evolved features that distinguish the group from related groups).[39] A 2011 study by Philip Mannion and Calvo found Andesauridae to be paraphyletic (excluding some of the group's descendants) and likewise recommended its disuse.[42]
In 2004, Upchurch and colleagues introduced a new group called Lithostrotia that included the more derived (evolved) members of Titanosauria. Argentinosaurus was classified outside this group and thus as a more basal ("primitive") titanosaurian.[32]: 278 The basal position within Titanosauria was confirmed by a number of subsequent studies.[38][30][43][44][45] In 2007, Calvo and colleagues named Futalognkosaurus; they found it to form a clade with Mendozasaurus and named it Lognkosauria.[46] A 2017 study by Carballido and colleagues recovered Argentinosaurus as a member of Lognkosauria and the sister taxon of Patagotitan.[8] In 2018, González Riga and colleagues also found it to belong in Lognkosauria, which in turn was found to belong to Lithostrotia.[47]
Another 2018 study by Hesham Sallam and colleagues found two different phylogenetic positions for Argentinosaurus based on two data sets. They did not recover it as a lognkosaurian but as either a basal titanosaur or a sister taxon of the more derived Epachthosaurus.[48] In 2019, Julian Silva Junior and colleagues found Argentinosaurus to belong to Lognkosauria once again; they recovered Lognkosauria and Rinconsauria (another group generally included in Titanosauria) to be outside Titanosauria.[49] Another 2019 study by González Riga and colleagues also found Argentinosaurus to belong to Lognkosauria; they found this group to form a larger clade with Rinconsauria within Titanosauria, which they named Colossosauria.[50]

|
Topology according to Carballido and colleagues, 2017.[8]
|
Topology according to González Riga and colleagues, 2019.[50]
|
Palaeobiology

The giant size of Argentinosaurus and other sauropods was likely made possible by a combination of factors; these include fast and energy-efficient feeding allowed for by the long neck and lack of mastication, fast growth and fast population recovery due to their many small offspring. Advantages of giant sizes would likely have included the ability to keep food inside the digestive tract for lengthy periods to extract a maximum of energy, and increased protection against predators.[51] Sauropods were oviparous (egg-laying). In 2016, Mark Hallett and Matthew Wedel stated that the eggs of Argentinosaurus were probably only 1 litre (0.26 US gal) in volume, and that a hatched Argentinosaurus was no longer than 1 metre (3.3 ft) and not heavier than 5 kilograms (11 lb). The largest sauropods increased their size by five orders of magnitude after hatching, more than in any other amniote animals.[52]: 186 Hallett and Wedel argued size increases in the evolution of sauropods were commonly followed by size increases of their predators, theropod dinosaurs. Argentinosaurus might have been preyed on by Mapusaurus, which is among the largest theropods known. Mapusaurus is known from at least seven individuals found together,[53] raising the possibility that this theropod hunted in packs to bring down large prey including Argentinosaurus.[52]: 206–207
In 2013, Sellers and colleagues used a computer model of the skeleton and muscles of Argentinosaurus to study its speed and gait. Before computer simulations, the only way of estimating speeds of dinosaurs was through studying anatomy and trackways. The computer model was based on a laser scan of a mounted skeletal reconstruction on display at the Museo Carmen Funes. Muscles and their properties were based on comparisons with living animals; the final model had a mass of 83 tonnes (91 short tons). Using computer simulation and machine learning techniques, which found a combination of movements that minimised energy requirements, the digital Argentinosaurus learned to walk. The optimal gait found by the algorithms was close to a pace (forelimb and hind limb on the same side of the body move simultaneously).[14] The model reached a top speed of just over 2 m/s (7.2 km/h, 5 mph).[54] The authors concluded with its giant size, Argentinosaurus reached a functional limit. Much larger terrestrial vertebrates might be possible but would require different body shapes and possibly behavioural change to prevent joint collapse. The authors of the study cautioned the model is not fully realistic and too simplistic, and that it could be improved in many areas. For further studies, more data from living animals is needed to improve the soft tissue reconstruction, and the model needs to be confirmed based on more complete sauropod specimens.[14]
Palaeoenvironment

Argentinosaurus was discovered in the Argentine Province of Neuquén. It was originally reported from the Huincul Group of the Río Limay Formation,[2] which have since become known as the Huincul Formation and the Río Limay Subgroup, the latter of which is a subdivision of the Neuquén Group. This unit is located in the Neuquén Basin in Patagonia. The Huincul Formation is composed of yellowish and greenish sandstones of fine-to-medium grain, some of which are tuffaceous.[55] These deposits were laid down during the Upper Cretaceous, either in the middle Cenomanian to early Turonian stages[56] or the early Turonian to late Santonian.[57] The deposits represent the drainage system of a braided river.[58]
Fossilised pollen indicates a wide variety of plants were present in the Huincul Formation. A study of the El Zampal section of the formation found hornworts, liverworts, ferns, Selaginellales, possible Noeggerathiales, gymnosperms (including gnetophytes and conifers), and angiosperms (flowering plants), in addition to several pollen grains of unknown affinities.[59] The Huincul Formation is among the richest Patagonian vertebrate associations, preserving fish including dipnoans and gar, chelid turtles, squamates, sphenodonts, neosuchian crocodilians, and a wide variety of dinosaurs.[56][60] Vertebrates are most commonly found in the lower, and therefore older, part of the formation.[61]
In addition to Argentinosaurus, the sauropods of the Huincul Formation are represented by another titanosaur, Choconsaurus,[62] and several rebbachisaurids including Cathartesaura,[63] Limaysaurus,[64][65] and some unnamed species.[61] Theropods including carcharodontosaurids such as Mapusaurus,[53] abelisaurids including Skorpiovenator,[66] Ilokelesia, and Tralkasaurus,[67] noasaurids such as Huinculsaurus,[68] paravians such as Overoraptor,[69] and other theropods such as Aoniraptor and Gualicho[70] have also been discovered there.[56] Several iguanodonts are also present in the Huincul Formation.[55]
References
- 1 2 3 Salgado, L.; Powell, J.E. (2010). "Reassessment of the vertebral laminae in some South American titanosaurian sauropods". Journal of Vertebrate Paleontology. 30 (6): 1760–1772. Bibcode:2010JVPal..30.1760S. doi:10.1080/02724634.2010.520783. hdl:11336/73562. S2CID 86501269.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Bonaparte, J.; Coria, R. (1993). "Un nuevo y gigantesco sauropodo titanosaurio de la Formacion Rio Limay (Albiano-Cenomaniano) de la Provincia del Neuquen, Argentina". Ameghiniana (in Spanish). 30 (3): 271–282.
- ↑ Montanaro, P. (February 3, 2019). "A 30 años del hallazgo del dino gigante de Huincul". Lmneuquen.com (in Spanish). Retrieved November 25, 2019.
- ↑ Prothero, D.R. (2016). Giants of the Lost World: Dinosaurs and Other Extinct Monsters of South America. Smithsonian Institution. ISBN 978-1-58834-574-5.
- 1 2 3 4 5 Mazzetta, G.V.; Christiansen, P.; Fariña, R.A. (2004). "Giants and bizarres: Body size of some southern South American Cretaceous dinosaurs" (PDF). Historical Biology. 16 (2–4): 71–83. Bibcode:2004HBio...16...71M. CiteSeerX 10.1.1.694.1650. doi:10.1080/08912960410001715132. S2CID 56028251. Archived (PDF) from the original on August 19, 2018.
- 1 2 3 Salgado, L.; Bonaparte, J.F. (2007). "Sauropodomorpha". In Gasparini, Z.; Salgado, L.; Coria, R.A. (eds.). Patagonian Mesozoic Reptiles. Bloomington and Indianapolis: Indiana University Press. pp. 188–228. ISBN 978-0-253-34857-9.
- 1 2 3 4 5 6 7 8 9 Paul, G.S. (2019). "Determining the largest known land animal: A critical comparison of differing methods for restoring the volume and mass of extinct animals" (PDF). Annals of the Carnegie Museum. 85 (4): 335–358. doi:10.2992/007.085.0403. S2CID 210840060.
- 1 2 3 4 5 Carballido, J.L.; Pol, D.; Otero, A.; Cerda, I.A.; Salgado, L.; Garrido, A.C.; Ramezani, J.; Cúneo, N.R.; Krause, J.M. (August 16, 2017). "A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs". Proceedings of the Royal Society B: Biological Sciences. 284 (1860): 20171219. doi:10.1098/rspb.2017.1219. PMC 5563814. PMID 28794222.
- ↑ Paul, G.S. (1997). "Dinosaur models: the good, the bad, and using them to estimate the mass of dinosaurs" (PDF). In Wolberg, D.L.; Stump, E.; Rosenberg, G.D. (eds.). DinoFest International Proceedings. Dinofest International. The Academy of Natural Sciences. pp. 129–154. Archived (PDF) from the original on March 4, 2016.
- 1 2 Paul, G.S. (1994). "Big sauropods – really, really big sauropods" (PDF). The Dinosaur Report: 12–13. Archived (PDF) from the original on March 6, 2012.
- ↑ Appenzeller, T. (1994). "Argentine dinos vie for heavyweight titles" (PDF). Science. 266 (5192): 1805. Bibcode:1994Sci...266.1805A. doi:10.1126/science.266.5192.1805. PMID 17737065. Archived (PDF) from the original on April 14, 2019. Retrieved November 20, 2019.
- 1 2 Carpenter, K. (2006). Foster, J.R.; Lucas, S.G. (eds.). "Biggest of the big: A critical re-evaluation of the mega-sauropod Amphicoelias fragillimus Cope, 1878" (PDF). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin. 36: 131–138. Archived from the original (PDF) on March 8, 2016. Retrieved December 9, 2016.
- 1 2 Calvo, J.O.; Juárez Valieri, R.D.; Porfiri, J.D. (2008). "Re-sizing giants: estimation of body lenght [sic] of Futalognkosaurus dukei and implications for giant titanosaurian sauropods". Congreso Latinoamericano de Paleontología de Vertebrados. Retrieved November 20, 2018.
- 1 2 3 4 Sellers, W.I.; Margetts, L.; Coria, R. A.B.; Manning, P.L. (2013). Carrier, D. (ed.). "March of the titans: The locomotor capabilities of sauropod dinosaurs". PLOS ONE. 8 (10): e78733. Bibcode:2013PLoSO...878733S. doi:10.1371/journal.pone.0078733. PMC 3864407. PMID 24348896.
- ↑ Hartman, S. (2013). "The biggest of the big". Skeletal Drawing. Archived from the original on November 5, 2018. Retrieved November 4, 2018.
- 1 2 Paul, G.S. (October 25, 2016). The Princeton Field Guide to Dinosaurs (2nd ed.). Princeton, N.J.: Princeton University Press. ISBN 978-0-691-16766-4. OCLC 954055249.
- ↑ Benson, R.B.J.; Campione, N.S.E.; Carrano, M.T.; Mannion, P.D.; Sullivan, C.; Upchurch, P.; Evans, D.C. (2014). "Rates of dinosaur body mass evolution indicate 170 million years of sustained ecological innovation on the avian stem lineage". PLOS Biology. 12 (5): e1001853. doi:10.1371/journal.pbio.1001853. PMC 4011683. PMID 24802911.
- ↑ Benson, R. B. J.; Hunt, G.; Carrano, M.T.; Campione, N.; Mannion, P. (2018). "Cope's rule and the adaptive landscape of dinosaur body size evolution". Palaeontology. 61 (1): 13–48. Bibcode:2018Palgy..61...13B. doi:10.1111/pala.12329.
- 1 2 Campione, Nicolás E.; Evans, David C. (2020). "The accuracy and precision of body mass estimation in non-avian dinosaurs". Biological Reviews. 95 (6): 1759–1797. doi:10.1111/brv.12638. ISSN 1469-185X. PMID 32869488. S2CID 221404013.
- ↑ González Riga, B.J.; Lamanna, M.C.; Ortiz David, L.D.; Calvo, J. O.; Coria, J.P. (2016). "A gigantic new dinosaur from Argentina and the evolution of the sauropod hind foot". Scientific Reports. 6: 19165. Bibcode:2016NatSR...619165G. doi:10.1038/srep19165. PMC 4725985. PMID 26777391.
- ↑ Paul, Gregory S.; Larramendi, Asier (June 9, 2023). "Body mass estimate of Bruhathkayosaurus and other fragmentary sauropod remains suggest the largest land animals were about as big as the greatest whales". Lethaia. 56 (2): 1–11. Bibcode:2023Letha..56..2.5P. doi:10.18261/let.56.2.5. S2CID 259782734.
- ↑ Lacovara, K.J.; Ibiricu, L.M.; Lamanna, M.C.; Poole, J.C.; Schroeter, E.R.; Ullmann, P.V.; Voegele, K.K.; Boles, Z.M.; Egerton, V.M.; Harris, J.D.; Martínez, R.D.; Novas, F.E. (September 4, 2014). "A gigantic, exceptionally complete titanosaurian sauropod dinosaur from Southern Patagonia, Argentina". Scientific Reports. 4: 6196. Bibcode:2014NatSR...4E6196L. doi:10.1038/srep06196. PMC 5385829. PMID 25186586.
- ↑ Fowler, D.W.; Sullivan, Robert M. (2011). "The first giant titanosaurian sauropod from the Upper Cretaceous of North America" (PDF). Acta Palaeontologica Polonica. 56 (4): 685–690. doi:10.4202/app.2010.0105. S2CID 53126360.
- ↑ Bates, K.T.; Falkingham, P.L.; Macaulay, S.; Brassey, C.; Maidment, S.C.R. (2015). "Downsizing a giant: re-evaluating Dreadnoughtus body mass". Biology Letters. 11 (6): 20150215. doi:10.1098/rsbl.2015.0215. ISSN 1744-9561. PMC 4528471. PMID 26063751.
- ↑ Tschopp, E.; Mateus, O.V.; Benson, R.B.J. (2015). "A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)". PeerJ. 3: e857. doi:10.7717/peerj.857. PMC 4393826. PMID 25870766.
- ↑ Lovelace, D.M.; Hartman, S.A.; Wahl, W.R. (2007). "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny". Arquivos do Museu Nacional. 65 (4): 527–544.
- ↑ "The life of the blue whale, Record breaker".
- ↑ Goldbogen, J. A.; Calambokidis, J.; Oleson, E.; Potvin, J.; Pyenson, N.D.; Schorr, G.; Shadwick, R.E. (2011). "Mechanics, hydrodynamics and energetics of blue whale lunge feeding: efficiency dependence on krill density". Journal of Experimental Biology. 214 (Pt 1): 131–146. doi:10.1242/jeb.048157. PMID 21147977. S2CID 12100333.
- 1 2 Bonaparte, J.F. (1996). "Cretaceous tetrapods of Argentina". Münchner Geowissenschaftliche Abhandlungen. 30: 73–130.
- 1 2 3 4 5 6 Novas, F.E. (2009). The Age of Dinosaurs in South America. Bloomington: Indiana University Press. pp. 204–205. ISBN 978-0-253-35289-7.
- ↑ Wedel, M.J. (2005). "Postcranial skeletal pneumaticity in sauropods and its implications for mass estimates". In Rogers, C.C.; Wilson, J.A. (eds.). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. pp. 201–228. ISBN 9780520246232.
- 1 2 3 Upchurch, P.; Barret, P.M.; Dodson, P. (2004). "Sauropoda". In Weishampel, D.B.; Dodson, P.; Osmólska, H. (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 259–322. ISBN 978-0-520-25408-4.
- ↑ Coria, R.A.; Filippi, L.S.; Chiappe, L.M.; García, R.; Arcucci, A.B. (2013). "Overosaurus paradasorum gen. et sp. nov. , a new sauropod dinosaur (Titanosauria: Lithostrotia) from the Late Cretaceous of Neuquén, Patagonia, Argentina". Zootaxa. 3683 (4): 357–376. doi:10.11646/zootaxa.3683.4.2. hdl:11336/21928. PMID 25250458.
- ↑ Novas, F.E.; Ezcurra, M. (2006). "Reinterpretation of the dorsal vertebrae of Argentinosaurus huinculensis (Sauropoda, Titanosauridae)". Ameghiniana. 43 (4): 48–49R.
- ↑ Sanz, J.L.; Powell, J.E.; Le Loeuff, J.; Martínez, R.; Pereda Suberbiola, X. (1999). "Sauropod remains from the Upper Cretaceous of Laño (northcentral Spain). Titanosaur phylogenetic relationships". Estudios del Museo de Ciencias Naturales de Alava. 14 (1): 235–255.
- ↑ Powell, J.E. (2003). Revision of South American Titanosaurid Dinosaurs: Palaeobiological, Palaeobiogeographical and Phylogenetic Aspects. Queen Victoria Museum and Art Gallery.
- ↑ Apesteguía, S. (2005). "Evolution of the hyposphene-hypantrum complex within Sauropoda". In Tidwell, V.; Carpenter, K. (eds.). Thunder-Lizards: The Sauropodomorph Dinosaurs. Bloomington and Indianapolis: Indiana University Press. ISBN 978-0-253-34542-4.
- 1 2 Wilson, J.A. (2006). "An overview of titanosaur evolution and phylogeny". Actas de las III Jornadas Sobre Dinosaurios y Su Entorno. Burgos: Salas de los Infantes. 169: 169–190.
- 1 2 Wilson, J.A.; Upchurch, P. (2003). "A revision of Titanosaurus Lydekker (Dinosauria - Sauropoda), the first dinosaur genus with a 'Gondwanan' distribution" (PDF). Journal of Systematic Palaeontology. 1 (3): 125–160. doi:10.1017/S1477201903001044. S2CID 53997295. Archived (PDF) from the original on May 14, 2016. Retrieved January 11, 2020.
- ↑ Salgado, L.; Coria, R.A.; Calvo, J.O. (1997). "Evolution of titanosaurid sauropods I.: Phylogenetic analysis based on the postcranial evidence". Ameghiniana. 34 (1): 3–32.
- ↑ Pisani, D.; Yates, A.M.; Langer, M.C.; Benson, M.J. (2002). "A genus-level supertree of the Dinosauria". Proceedings of the Royal Society of London. Series B: Biological Sciences. 269 (1494): 915–921. doi:10.1098/rspb.2001.1942. PMC 1690971. PMID 12028774.
- ↑ Mannion, P.D.; Calvo, J.O. (2011). "Anatomy of the basal titanosaur (Dinosauria, Sauropoda) Andesaurus delgadoi from the mid-Cretaceous (Albian–early Cenomanian) Río Limay Formation, Neuquén Province, Argentina: implications for titanosaur systematics". Zoological Journal of the Linnean Society. 163 (1): 155–181. doi:10.1111/j.1096-3642.2011.00699.x.
- ↑ Filippi, L.S.; García, R.A.; Garrido, A.C. (2011). "A new titanosaur sauropod dinosaur from the Upper Cretaceous ofNorth Patagonia, Argentina" (PDF). Acta Palaeontologica Polonica. 56 (3): 505–520. doi:10.4202/app.2010.0019. S2CID 62837802. Archived (PDF) from the original on April 17, 2018. Retrieved November 14, 2019.
- ↑ Lacovara, K.J.; Ibiricu, L.M.; Lamanna, M.C.; Poole, J.C.; Schroeter, E.R.; Ullmann, P.V.; Voegele, K.K.; Boles, Z.M.; Egerton, V.M.; Harris, J.D.; Martínez, R.D.; Novas, F.E. (September 4, 2014). "A Gigantic, Exceptionally Complete Titanosaurian Sauropod Dinosaur from Southern Patagonia, Argentina". Scientific Reports. 4: 6196. Bibcode:2014NatSR...4E6196L. doi:10.1038/srep06196. PMC 5385829. PMID 25186586.
- ↑ González Riga, B.J.; Lamanna, M.C.; Ortiz David, L.D.; Calvo, J.O.; Coria, J.P. (2016). "A gigantic new dinosaur from Argentina and the evolution of the sauropod hind foot". Scientific Reports. 6: 19165. Bibcode:2016NatSR...619165G. doi:10.1038/srep19165. ISSN 2045-2322. PMC 4725985. PMID 26777391.
- ↑ Calvo, Jorge O.; Porfiri, Juan D.; González-Riga, Bernardo J.; Kellner, A.W.A. (2007). "A new Cretaceous terrestrial ecosystem from Gondwana with the description of a new sauropod dinosaur". Anais da Academia Brasileira de Ciências. 79 (3): 529–541. doi:10.1590/S0001-37652007000300013. PMID 17768539.
- ↑ Gonzalez Riga, B.J.; Mannion, P.D.; Poropat, S.F.; Ortiz David, L.; Coria, J.P. (2018). "Osteology of the Late Cretaceous Argentinean sauropod dinosaur Mendozasaurus neguyelap: implications for basal titanosaur relationships" (PDF). Journal of the Linnean Society. 184 (1): 136–181. doi:10.1093/zoolinnean/zlx103. hdl:10044/1/53967. Archived (PDF) from the original on April 28, 2019. Retrieved September 23, 2019.
- ↑ Sallam, H.M.; Gorscak, E.; O'Connor, P.M.; El-Dawoudi, I.M.; El-Sayed, S.; Saber, S.; Kora, M.A.; Sertich, J.J.W.; Seiffer, E.R.; Lamanna, M.C. (2018). "New Egyptian sauropod reveals Late Cretaceous dinosaur dispersal between Europe and Africa" (PDF). Nature Ecology & Evolution. 2 (3): 445–451. Bibcode:2018NatEE...2..445S. doi:10.1038/s41559-017-0455-5. PMID 29379183. S2CID 3375335. Archived (PDF) from the original on August 28, 2019. Retrieved October 30, 2019.
- ↑ Silva Junior, J.C.G.; Marinho, T.S.; Martinelli, A.G.; Langer, M.C. (2019). "Osteology and systematics of Uberabatitan ribeiroi (Dinosauria; Sauropoda): a Late Cretaceous titanosaur from Minas Gerais, Brazil". Zootaxa. 4577 (3): 401–438. doi:10.11646/zootaxa.4577.3.1. PMID 31715707. S2CID 145939866.
- 1 2 González Riga, B.J.; Lamanna, M.C.; Otero, A.; Ortiz David, L.D.; Kellner, A.W.A.; Ibiricu, L.M. (2019). "An overview of the appendicular skeletal anatomy of South American titanosaurian sauropods, with definition of a newly recognized clade". Anais da Academia Brasileira de Ciências. 91 (suppl 2): e20180374. doi:10.1590/0001-3765201920180374. hdl:11336/106658. PMID 31340217.
- ↑ Sander, P.M.; Christian, A.; Clauss, M.; Fechner, R.; Gee, C.T.; Griebeler, E.M.; Gunga, Hanns-Christian; Hummel, J.; Mallison, H.; Perry, S.F. (2011). "Biology of the sauropod dinosaurs: the evolution of gigantism". Biological Reviews. 86 (1): 117–155. doi:10.1111/j.1469-185X.2010.00137.x. PMC 3045712. PMID 21251189.
- 1 2 Hallett, M.; Wedel, M. (2016), The Sauropod Dinosaurs: Life in the Age of Giants, Baltimore: Johns Hopkins University Press, ISBN 978-1421420288
- 1 2 Coria, R.A.; Currie, P.J. (2006). "A new carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina". Geodiversitas. 28 (1): 71–11.
- ↑ "Scientists digitally reconstruct giant steps taken by dinosaurs". The University of Manchester. October 30, 2013. Archived from the original on December 29, 2019. Retrieved December 29, 2019.
- 1 2 Leanza, H.A; Apesteguı́a, S.; Novas, F.E; de la Fuente, M.S. (February 1, 2004). "Cretaceous terrestrial beds from the Neuquén Basin (Argentina) and their tetrapod assemblages". Cretaceous Research. 25 (1): 61–87. Bibcode:2004CrRes..25...61L. doi:10.1016/j.cretres.2003.10.005. ISSN 0195-6671.
- 1 2 3 Motta, M.J.; Aranciaga Rolando, A.M.; Rozadilla, S.; Agnolín, F.E.; Chimento, N.R.; Egli, F.B.; Novas, F.E. (2016). "New theropod fauna from the upper cretaceous (Huincul Formation) of Northwestern Patagonia, Argentina". New Mexico Museum of Natural History and Science Bulletin. 71: 231–253.
- ↑ Corbella, H.; Novas, F.E.; Apesteguía, S.; Leanza, H. (2004). "First fission-track age for the dinosaur-bearing Neuquén Group (Upper Cretaceous), Neuquén Basin, Argentina". Revista del Museo Argentino de Ciencias Naturales. Nueva Serie. 6 (21): 227=232. doi:10.22179/REVMACN.6.84.
- ↑ Rainoldi, A.L.; Franchini, Marta; Beaufort, D.; Mozley, P.; Giusiano, A.; Nora, C.; Patrier, P.; Impiccini, A.; Pons, J. (2015). "Mineral reactions associated with hydrocarbon paleomigration in the Huincul High, Neuquén Basin, Argentina". GSA Bulletin. 127 (11–12): 1711–1729. Bibcode:2015GSAB..127.1711R. doi:10.1130/B31201.1. hdl:11336/36686.
- ↑ Vallati, P. (2001). "Middle cretaceous microflora from the Huincul Formation ("Dinosaurian Beds") in the Neuquén Basin, Patagonia, Argentina". Palynology. 25 (1): 179–197. Bibcode:2001Paly...25..179V. doi:10.2113/0250179.
- ↑ Motta, M.J.; Brissón Egli, F.; Aranciaga Rolando, A.M.; Rozadilla, S.; Gentil, A. R.; Lio, G.; Cerroni, M.; Garcia Marsà, J.; Agnolín, F. L.; D'Angelo, J. S.; Álvarez-Herrera, G. P.; Alsina, C.H.; Novas, F.E. (2019). "New vertebrate remains from the Huincul Formation (Cenomanian–Turonian;Upper Cretaceous) in Río Negro, Argentina". Publicación Electrónica de la Asociación Paleontológica Argentina. 19 (1): R26. doi:10.5710/PEAPA.15.04.2019.295. hdl:11336/161858. S2CID 127726069. Archived from the original on December 14, 2019. Retrieved December 14, 2019.
- 1 2 Bellardini, F.; Filippi, L.S. (2018). "New evidence of saurischian dinosaurs from the upper member of the Huincul Formation (Cenomanian) of Neuquén Province, Patagonia, Argentina". Reunión de Comunicaciones de la Asociación Paleontológica Argentina: 10.
- ↑ Simón, E.; Salgado, L.; Calvo, J.O. (2017). "A new titanosaur sauropod from the Upper Cretaceous of Patagonia, Neuquén Province, Argentina". Ameghiniana. 55 (1): 1–29. doi:10.5710/AMGH.01.08.2017.3051. S2CID 134332465.
- ↑ de Jesus Faria, C.C.; Riga, B.G.; dos Anjos Candeiro, C.R.; da Silva Marinho, T.; David, L.O.; Simbras, F.M.; Castanho, R.B.; Muniz, F.P.; Gomes da Costa Pereira, P.V.L. (August 1, 2015). "Cretaceous sauropod diversity and taxonomic succession in South America". Journal of South American Earth Sciences. 61: 154–163. Bibcode:2015JSAES..61..154D. doi:10.1016/j.jsames.2014.11.008. hdl:11336/37899. ISSN 0895-9811.
- ↑ Calvo, J.O.; Salgado, L. (1995). "Rebbachisaurus tessonei sp. nov. a new Sauropoda from the Albian-Cenomanian of Argentina; new evidence on the origin of the Diplodocidae". Gaia. 11: 13–33.
- ↑ Salgado, L.; Garrido, A.; Cocca, S.E.; Cocca, J.R. (2004). "Lower Cretaceous rebbachisaurid sauropods from Cerro Aguada del León (Lohan Cura Formation), Neuquén Province, northwestern Patagonia, Argentina". Journal of Vertebrate Paleontology. 24 (4): 903–912. doi:10.1671/0272-4634(2004)024[0903:lcrsfc]2.0.co;2. S2CID 129233849.
- ↑ Canale, J.I.; Scanferla, C.A.; Agnolin, F.L.; Novas, F.E. (2009). "New carnivorous dinosaur from the Late Cretaceous of NW Patagonia and the evolution of abelisaurid theropods". Naturwissenschaften. 96 (3): 409–14. Bibcode:2009NW.....96..409C. doi:10.1007/s00114-008-0487-4. hdl:11336/52024. PMID 19057888. S2CID 23619863.
- ↑ Cerroni, M.A.; Motta, M.J.; Agnolín, F.L.; Aranciaga Rolando, A.M.; Brissón Egliab, F.; Novas, F.E. (2020). "A new abelisaurid from the Huincul Formation (Cenomanian-Turonian; Upper Cretaceous) of Río Negro province, Argentina". Journal of South American Earth Sciences. 98: 102445. Bibcode:2020JSAES..9802445C. doi:10.1016/j.jsames.2019.102445. S2CID 213781725.
- ↑ Baiano, M.A.; Coria, R.A.; Cau, A. (2020). "A new abelisauroid (Dinosauria: Theropoda) from the Huincul Formation (lower Upper Cretaceous, Neuquén Basin) of Patagonia, Argentina". Cretaceous Research. 110: 104408. Bibcode:2020CrRes.11004408B. doi:10.1016/j.cretres.2020.104408. S2CID 214118853.
- ↑ Matías J. Motta; Federico L. Agnolín; Federico Brissón Egli; Fernando E. Novas (2020). "New theropod dinosaur from the Upper Cretaceous of Patagonia sheds light on the paravian radiation in Gondwana". The Science of Nature. 107 (3): Article number 24. Bibcode:2020SciNa.107...24M. doi:10.1007/s00114-020-01682-1. hdl:11336/135530. PMID 32468191. S2CID 218913199.
- ↑ Apesteguía, S.; Smith, N.D.; Juárez Valieri, R.; Makovicky, P.J. (2016). "An unusual new theropod with a didactyl manus from the Upper Cretaceous of Patagonia, Argentina". PLOS ONE. 11 (7): e0157793. Bibcode:2016PLoSO..1157793A. doi:10.1371/journal.pone.0157793. PMC 4943716. PMID 27410683.
External links
 Media related to Argentinosaurus at Wikimedia Commons
Media related to Argentinosaurus at Wikimedia Commons